En un sentido amplio, metabolismo es el conjunto de todas las reacciones químicas que se producen en el interior de las células de un organismo. Mediante esas reacciones se transforman las moléculas nutritivas que, digeridas y transportadas por la sangre, llegan a ellas. Estas reacciones químicas metabólicas (repetimos, ambas reacciones suceden en las células) pueden ser de dos tipos: catabolismo y anabolismo .
La fermentación alcohólica, también conocida como, fermentación etílica, o del etanol, es un proceso de tipo biológico, en el cual se lleva a cabo una fermentación sin presencia de oxígeno. Este tipo de fermentación se debe a las actividades de ciertos microorganismos, los cuales se encargan de procesar azúcares, como la glucosa, la fructosa, etc. (hidratos de carbono), dando como resultado un alcohol a modo de etanol, CO2 (gas) y ATP (adenosín trifosfato), moléculas que son utilizadas por los propios microorganismos en sus metabolismos energéticos.
Numerosos hongos, bacterias, algas y algunos protozoos, fermentan azúcares, transformándolos en etanol y CO2. Este es el proceso que se conoce como fermentación alcohólica.En este tipo de fermentaciones, el piruvato (anión delácido piruvico, es descarboxilado, convirtiéndose en acetaldehído, el cual a su vez, es reducido a etanol a través de la enzima, alcohol deshidrogenasa, utilizando como dador de electrones al NADH..
La fermentación alcohólica es utilizada desde antiguo para realizar productos como la cerveza o el vino. Los griegos otorgaban el descubrimiento de este proceso al dios Dionisio. Y algunos procesos similares, como la destilación de alcohol, se llevaban a cabo ya en el año 1150. Sin duda, dichos procesos fueron esenciales para el desarrollo de la alquimia en la Edad Media. Los descubrimientos químicos posteriores llevaron al investigador, Gay-Lussac, a describir la reacción de fermentación que tenía lugar partiendo de la glucosa, con obtención de etanol, a pesar de que, por aquel entonces, aún no se conocía la fermentación alcohólica y sus fundamentos.
Fueron muchos científicos los que intentaron dar explicación al proceso que hoy conocemos como fermentación, pero hasta 1818 no se descubre que las causantes del proceso eran las levaduras. Pocos años después, se descubre la enzima responsable del proceso, la zimasa, otorgándose el Premio Nobel de Química en 1897, por dicho descubrimiento esencial, a Eduard Buchner. En los años posteriores, se siguió trabajando en el tema, hasta que en 1929, se descubre el cofactor NADH, esencial en el proceso de fermentación, pues su principal función es el intercambio de electrones.
Necesitamos la energía para realizar nuestras necesidades y actividades diarias.
La vía aeróbica. En esta vía aparece el oxígeno y utiliza el glucógeno, la glucosa y los ácidos grasos como la principal fuente para la producción de energía. La glucosa, procedente de la degradación del glucógeno, se oxidará a través del proceso de la glucólisis.
Por otra parte, los ácidos grasos se mezclarán con la Coenzima A, para posteriormente acceder al interior de la mitocondria. Para producirse esto, el ácido graso ayudado de la Coenzima A tendrán que unirse a la L-carnitina. La L-carnitina se encarga del transporte de los ácidos grasos al interior de la mitocondria, para producirse el acetil-CoA
Ácido graso + Coenzima A = AcilCoA
AcilCoa + L-carnitina = acetil-CoA
No solo participan en la obtención de energía por la vía aeróbica el glucógeno, la glucosa y los ácidos grasos, también podemos destacar que algunos aminoácidos, cetoácidos y glicerol pueden oxidarse para formar acetil-CoA o para formar glucosa (gluconeogénesis).
En el interior de la mitocondria, el acetil-CoA sufre un proceso de oxidación apoyado en el ciclo de Krebs. A través del proceso de Krebs se producen una serie de reacciones químicas que dan como resultado ATP, éste se encarga de convertir la energía química en energía mecánica.
La vía aeróbica entra en acción cuando los esfuerzos no son muy fuertes pero sí de una duración notable. Se calcula que se empieza a quemar grasa a los 25-30 minutos realizando una actividad física moderada sin realizar pausas.
En el músculo existen dos tipos de fibras: las fibras de tipo I, que cuentan con una gran capacidad del ejercicio aeróbico, y las fibras de tipo II, se contraen con una mayor rapidez.
Orgánulo citoplasmático de las células
eucariotas, de forma ovoidal,
formado por una doble membrana, que tiene como principal función la producción
de energía mediante el consumo de oxigeno, y la producción de dióxido de
carbono y agua como productos de la respiración celular.
Función de la Mitocondria
Membrana o Compartimiento
Funciones
Metabólicas
Membrana Externa
üSíntesis
de fosfolípidos.
üInsaturación de ácidos grasos.
üElongación de ácidos grasos.
Membrana Interna
üTransporte
de electrones.
üFosforilación oxidativa.
üTransporte
metabólico.
Matriz
üOxidación del piruvato.
üCiclo del TCA (ácido tricarboxílico).
üB-oxidación de lópidos.
üReplicación del DNA.
üSíntesis de RNA (transcripción).
üSíntesis de proteínas (traducción).
Membrana o Compartimiento
Funciones
Metabólicas
Estructura
Membranas
La mitocondria están rodeadas por dos membranas diferentes en lo que corresponde a función y actividad
enzimática. Éstas separan tres espacios: el citosol, el espacio intermembrana y la matriz mitocondrial.
Membrana externa: bicapa lipídica exterior la cual es permeable a iones, metabolitos y otras macromoléculas. Esto
último se debe a que contiene unas proteínas llamadas porinas, las cuales forman poros. Esta membrana realiza
relativamente pocas funciones enzimáticas o de transporte.
Membrana interna:ésta membrana contiene más proteínas, carece de poros y es más selectiva en comparación con la
membrana externa. Contiene muchos complejos enzimáticos y sistemas de transporte transmembrana; éstos están
implicados en la traslocación de moléculas.
Espacio intermembranoso:
Es un líquido similar al hialoplasma, este se localiza entre las dos membranas. Posee una alta concentración de
protones debido al bombeo de los mismos por los complejos enzimáticos de la cadena respiratoria. En este espacio se
encuentran diversas enzimas que intervienen en la trasferencia del enlace de alta energía del ATP.
Matriz mitocondrial:Contiene menos moléculas que el citosol, aunque tiene iones, metabolitos, ADN circular bicatenario, ribosomas
tipo 70S y contiene ARNm. Se podría decir que tiene todos los organelos que tendría una célula procariota. En ésta parte
de la mitocondria hay diversas rutas metabólicas esenciales para la vida, cómo el cíclo de Krebs y la beta-oxidación de
ácidos grasos, además de la oxidación de aminoácidos.
Proceso intramitocondrial e irreversible,en el cual el piruvato procedente de la glicolisis se oxida para dar lugar a acetil- CoA y CO2, mediante la accion del complejo de la piruvato DH.
El complejo Piruvato DH consta de tres enzimas:
-E1,piruvato DH, necesita como coenzima TPP.
-E2, dihidrolipiol transacetilasa, usa como coenzima lipoamida y traslada la Co-A.
-E3, dihidrolipoil DH, utiliza como coenzima FAD y modifica al NAD.
En resumen, esta formado por 3 enzimas y necesita 5 coenzimas.
1ºSe une el piruvato a la TPP de E1 formando hidroxietil-TPP y libera CO2.
2ºTralada el hidroxietilo al la lipoamida reduciendola y libera la TPP.
3ºTraslado del grupo acetil al CoA y libera la lipoamida reducida.
4º Se transfieren dos electrones de la lipoamida a FAD que se reduce a FADH2
5º El NAD oxida a FADH2 y se forma NADH que se dirige a cadena respiratoria.
Regulacion del proceso:
Modificacion alosterica del complejo PDH: ac.grasos de cadena larga, ATP, AMP, CoA, NADH son efectores negativos.
Cuando el ciclo de Krebs no dispone de suficiente acetil-CoA se activa alostericamente el complejo de piruvato DH.
Modificacion covalente del complejo PDH: a)por fosforilacion: la cinasa inhibe el complejo, se activa por incremento de acetil-CoA, NADH, ATP.
b)por desfosforilacion: la fosfatasa activa el complejo PDH, se activa por insulina.
Cadena de transporte de electrones
Es uno de los sistemas celulares más importantes. Se encuentra tanto en procariotas como en eucariotas. Este
hecho no solo resalta su importancia metabólica sino que, además ésta se ve corroborada por la poca alteración de las
proteínas que la componen a lo largo de la evolución. En los procariotas se encuentra adosado a la membrana plasmática y
en eucariotas las proteínas que forman la cadena de transporte de electrones se encuentran en las membranas internas de
cloroplastos y mitocondrias. De los tres la cadena transportadora de electrones de mitocondrias es la más conocida.
Complejo I
El complejo I o NADH deshidrogenasa o NADH: ubiquinona oxidoreductasa, capta dos electrones del NADH y
los transfiere a un transportador liposoluble denominado ubiquinona (Q). El producto reducido, que se conoce con el
nombre de ubiquinol (QH2) puede difundir libremente por la membrana. Al mismo tiempo, el Complejo I transloca cuatro
protones a través de membrana y produce un gradiente de protones.
Complejo II
El Complejo II o succinato deshidrogenasa; no es una bomba de protones. Además es la única enzima del ciclo de
Krebs asociado a membrana. Antes de que este complejo actúe el FADH2 se forma durante la conversión de succinato en
fumarato en el ciclo del ácido cítrico. A continuación los electrones son transferidos por medio de una serie de centros
FeS hacia Q. EL glicerol-3-fosfato y el acetil-CoA también transfieren electrones a Q mediante vías diferentes en que
participan flavoproteínas.
Complejo III
El complejo III o complejo citocromo bc1; obtiene dos electrones desde QH2 y los transfiere a dos moléculas de
citocromo c, que es un transportador de electrones hidrosoluble que se encuentra en el espacio intermembrana de la
mitocondria. Al mismo tiempo, transloca cuatro protones a través de la membrana por los dos electrones transportados
desde el ubiquinol.
Complejo IV
El complejo IV o citocromo c oxidasa; capta cuatro electrones de las cuatro moléculas de citocromo c y se
transfieren al oxígeno (O2), para producir dos moléculas de agua (H2O). Al mismo tiempo, se translocan cuatro protones
al espacio intermembrana, por los cuatro electrones. Además "desaparecen" de la matriz 2 protones que forman parte del
H2O.
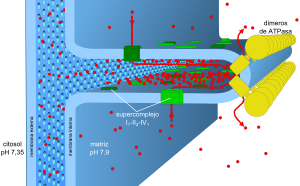
La fosforilación oxidativa es un proceso metabólico que utiliza energía liberada por la oxidación de nutrientes para producir adenosina trifosfato (ATP). Se le llama así para distinguirla de otras rutas que producen ATP con menor rendimiento, llamadas "a nivel de sustrato". Se calcula que hasta el 90% de la energía celular en forma de ATP es producida de esta forma.
Consta de dos etapas: en la primera, la energía libregenerada mediantereacciones químicas redoxen varioscomplejos multiproteicos-conocidos en su conjunto comocadena de transporte de electrones- se emplea para producir, por diversos procedimientos como bombeo, ciclos quinona/quinol o bucles redox, ungradiente electroquímicodeprotonesa través de unamembranaasociada en un proceso llamadoquimiosmosis. Lacadena respiratoriaestá formada por tres complejos de proteínas principales (complejo I,III,IV), y varios complejos "auxiliares", utilizando una variedad de donantes y aceptores deelectrones. Los tres complejos se asocian en supercomplejos para canalizar las moléculas transportadoras de electrones, lacoenzima Qy elcitocromo c, haciendo más eficiente el proceso.
La energía potencial de ese gradiente, llamada fuerza protón-motriz, se libera cuando se translocan los protones a través de un canal pasivo, la enzimaATP sintasa, y se utiliza en la adición de un grupo fosfato a una molécula de ADP para almacenar parte de esa energía potencial en los enlaces anhidro "de alta energía" de la molécula de ATP mediante un mecanismo en el que interviene la rotación de una parte de la enzima a medida que fluyen los protones a través de ella. En vertebrados, y posiblemente en todo el reino animal, se genera un ATP por cada 2,7 protones translocados. Algunos organismos tienen ATPasas con un rendimiento menor.
Existen también proteínas desacopladoras que permiten controlar el flujo de protones y generar calor desacoplando ambas fases de la fosforilación oxidativa.
Aunque las diversas formas de vida utilizan una gran variedad de nutrientes, casi todas realizan la fosforilación oxidativa para producir ATP, la molécula que provee de energía al metabolismo. Esta ruta es tan ubicua debido a que es una forma altamente eficaz de liberación de energía, en comparación con los procesos alternativos de fermentación, como la glucólisis anaeróbica.
Pese a que la fosforilación oxidativa es una parte vital del metabolismo, produce una pequeña proporción de especies reactivas del oxígenotales comosuperóxidoyperóxido de hidrógeno, lo que lleva a la propagación deradicales libres, provocando daño celular, contribuyendo aenfermedadesy, posiblemente, alenvejecimiento. Sin embargo, los radicales tienen un importante papel en laseñalización celular, y posiblemente en la formación de enlaces disulfuro de las propias proteínas de la membrana interna mitocondrial. Las enzimas que llevan a cabo esta ruta metabólica son blanco de muchas drogas y productos tóxicos queinhibensu actividad.
Esquema actual del sistema mitocondrial de la fosforilación oxidativa. Los equivalentes reducidos que se generan en el metabolismo (NADH, FADH2) son ácidos oxidados por la cadena de transporte de electrones. La energía libre generada en estareacción se emplea para bombearprotones (puntos rojos) desde la matriz mitocondrial hasta el interior de las crestas mitocondriales, para dar lugar a la fuerza protón-motriz. Cuando éste se disipa a través del retorno a la matriz de los protones a través de la ATP sintasa, la energía almacenada se emplea parafosforilar el ADP con un grupo fosfato para formar ATP.
La fosforilación oxidativa funciona con dos tipos de reacciones que están acopladas, una utiliza reacciones químicas que liberan energía, mientras que la otra utiliza esa energía para llevar a cabo sus reacciones. El flujo de electrones a través de la cadena de transporte de electrones, desde donantes de electrones como NADH a aceptores de electrones tales como oxígeno, es un proceso exergónico y libera energía, mientras que la síntesis de ATP es un proceso endergónico, el cual requiere de energía. Tanto la cadena de transporte de electrones como la ATP sintasa, están embebidos en la membrana, y la energía es transferida de la cadena de transporte de electrones a la ATP sintasa por el movimiento de protones a través de la membrana, en un proceso llamado quimiosmosis.
sábado, 25 de febrero de 2017
Quimiosmosis
Es la difusión de iones a través de una membrana. Específicamente, se relaciona con la generación de ATP mediante el movimiento de iones hidrógeno (protones o H+) a través de la membrana interna mitocondrial y de la membrana de los tilacoides de los cloroplastos.
Los protones difunden desde un área de alta concentración a un área de baja concentración. Peter Mitchell propuso que un gradiente de concentración electroquímico de protones a través de la membrana podía ser usado para crear ATP. Él vio un paralelismo con el proceso de ósmosis (difusión de agua a través de una membrana) y por esto fue denominado "quimiosmosis".
La ATP-sintasa es la enzima que produce ATP por quimiosmosis. Permite el paso de protones a través de ella, utilizando esa energía cinética para fosforilar ADP y así crear ATP. La generación de ATP por quimiosmosis ocurre en cloroplastos y mitocondrias, como también en algunas bacterias.
Peter Mitchell propuso la "hipótesis quimiosmótica" en 1961.1 Esta teoría propone esencialmente que la mayor parte de la síntesis de ATP en la respiración celular, viene de un gradiente electroquímico existente entre la membrana interna y el espacio intermembrana de la mitocondria, mediante el uso de la energía de NADH y FADH2 que se han formado por la ruptura de moléculas ricas en energía, como la glucosa.
Los transportadores traspasan electrones a la cadena transportadora de electrones en la membrana mitocondrial interna, que luego los traspasan a otras proteínas en la cadena transportadora. La energía disponible en los electrones se usa para bombear protones desde la matriz, a través de la membrana mitocondrial interna, guardando energía en forma de un gradiente electroquímico transmembrana. Los protones se devuelven a través de la membrana interna, mediante la enzima ATP-sintasa. El flujo de protones de vuelta a la matriz mitocondrial mediante la ATP-sintasa, provee de suficiente energía para que el ADP se combine con fósforo inorgánico para formar ATP. Los electrones y protones en la última bomba proteica de la cadena transportadora son llevados al oxígeno (O2) para formar agua (H2O).Las moléculas como la glucosa, son metabolizadas para producir acetil-CoA como un intermediario rico en energía. La oxidación de acetil-CoA en la matriz mitocondrial está acoplada a la reducción de una molécula transportadora como NAD+ y FAD.2
Ésta fue una propuesta radical en su tiempo, y no fue bien aceptada. La visión que prevalecía era que la energía de la transferencia electrónica se almacenaba es un intermediario estable de alta energía, un concepto mas conservativo del punto de vista químico. El problema de éste viejo paradigma fue que nunca se encontró aquel intermediario, y la evidencia del bombeo de protones por los complejos de la Cadena de Transporte de electrones creció de forma tal, que no pudo ser ignorada. Eventualmente, el peso de la evidencia comenzó a favorecer la hipótesis quimiosmótica, y en 1978, el Premio Nobel de química fue entregado a Peter Mitchell.3
El acoplamiento quimiosmótico es importante en la producción de ATP en el cloroplasto4 y muchos tipos de bacteria.5
La quimiosmosis se puede prrsentar en diferentes organismos como:
Quimiosmosis en mitocondrias
La rotura completa de una molécula de glucosa en presencia de oxígeno es denominada respiración celular. Las últimas etapas de éste proceso ocurren en la mitocondria. Las moléculas de alta energía NADH y FADH2 -generadas por el ciclo de Krebs- liberan los electrones hacia una cadena transportadora de electrones para crear una gradiente de protones a través de la membrana interna mitocondrial. La ATP-sintasa es luego usado para generar ATP por quimiosmosis. Éste proceso se conoce como fosforilación oxidativa porque el oxígeno es el último aceptor electrónico en la cadena transportadora mitocondrial.
La fosforilación quimiosmótica es la tercera y final vía biológica responsable por la producción de ATP mediante fosfato inorgánico y ADP a través de la fosforilación oxidativa.
Ocurriendo en la mitocondria de las células, la energía química de NADH -producido por el ciclo de Krebs- es utilizada para construir un gradiente de iones de hidrógeno (protones) con una concentración mayor en las crestas mitocondriales y en menor concentración en la matriz mitocondrial. Éste es el único paso de la fosforilación oxidativa que requiere de oxígeno: éste es utilizado como aceptor de electrones, combinándose con electrones libres e iones de hidrógeno para formar agua.
Quimiosmosis en plantas
Las reacciones luz-dependientes de la fotosíntesis, generan energía mediante quimiosmosis. La clorofila pierde un par de electrones al ser excitada o energizada por la luz. Este electrón viaja a través de una cadena transportadora de electrones, terminando parte de NADPH, una molécula de alta energía. El gradiente electroquímico generado a través de la membrana del tilacoide conduce a la producción de ATP mediante la ATP-sintasa. Este proceso se conoce como fotofosforilación.
Quimiosmosis en bacterias
Las bacterias también pueden utilizar la quimiosmosis para generar ATP. Las cianobacterias, bacterias verdes del azufre y bacterias púrpuras crean energía por un proceso llamado fotofosforilación. Estas bacterias usan la energía de la luz para crear un gradiente de protones usando una cadena trasportadora de electrones fotosintética. Algunas bacterias no-fotosintetizadoras como la E. coli, también contiene ATP-sintasa. De hecho, se cree que las mitocondrias y los cloroplastos se formaron cuando las células eucariontes tempranas ingirieron bacterias que pudieran crear energía mediante la quimiosmosis. Esto es denominado teoría endosimbiótica.